Diving
Inside this section
• Comparing limits
• The dive pattern
• Why dive so deep?
• Record dives by air-breathers
• Human diving problems
________________________________
Comparing limits with humans

Sources of data cited with Dive Stats table below.



The distinctive dive pattern

See a dive pattern for a juvenile elephant seal that was tracked by Jessica Meir. Although the exact path isn't known, this animal crossed Monterey Bay from south to north.

Why dive so deep?
Access to more ocean - With or without scuba gear, human divers barely scrape the surface of the ocean. Wide areas of the world oceans are 3,000-6,000 meters deep and exceptionally deep areas occur in narrow trenches more than 11,000 meters deep.
• Vertical migration
• FAO - Midwater trawling
What is so interesting down there? During the day, around the world's open oceans, many prey species congregate in one or more layers at midwater depths near 100-500 meters. These daytime aggregations can be so thick that sonar waves bounce off these layers, as if they were solid like the ground. This daytime feature is called the deep-scattering layer or DSL. At night, the DSL disappears as these animals spread out and move closer to the surface to hunt for surface-dwellers under the cloak of darkness. This daily mass movement of life, called the diel vertical migration is the largest multispecies migration on the planet and it happens every sunset and sundown as the deep scattering layers break up and reform.
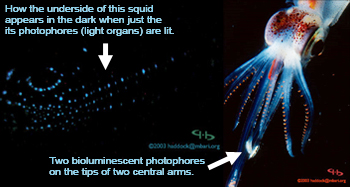 Bioluminescence in a small squid in the genus Abraliopsis.
If it's totally dark at depth, how do elephant seals home in on their prey and why do they need good eyesight? It's not totally dark above 200 meters depth so prey can be seen silhouetted against an upward view. Many animals that live at depth can also be spotted by their bioluminescence.
Photographer - Steven Haddock.The Bioluminescence Web Page.
Bioluminescence in a small squid in the genus Abraliopsis.
If it's totally dark at depth, how do elephant seals home in on their prey and why do they need good eyesight? It's not totally dark above 200 meters depth so prey can be seen silhouetted against an upward view. Many animals that live at depth can also be spotted by their bioluminescence.
Photographer - Steven Haddock.The Bioluminescence Web Page.
Most air-breathing animals can't get to the depths where all this tasty seafood hides during the day. By being able to dive deep, elephant seals are able to leave behind competitors who might outcompete them back at the surface and to avoid shallow-roaming large predators that want to eat them.
Elephant seals are specially outfitted to hunt in this environment where it is deep, cold and dark. Blubber, big eyes with keen vision that can sense flashes of bioluminescence in darkness, and many other adaptations make their lifestyle possible.
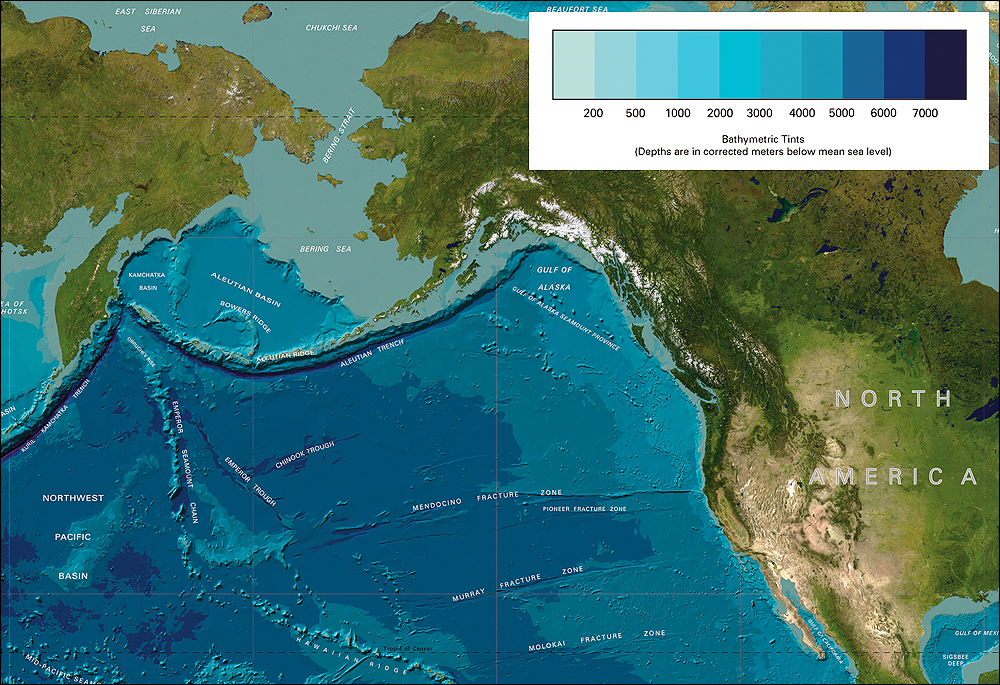 Bathymetric map of the N. Pacific.
Click on image to view larger map.
Image reproduced from the GEBCO world map, http://www.gebco.net.
Bathymetric map of the N. Pacific.
Click on image to view larger map.
Image reproduced from the GEBCO world map, http://www.gebco.net.
Avoiding coastal predators
Killer whales
Great white sharks |
 Killer whales belonging to "L-pod" just offshore Monterey, California, March 25, 2009. Photographer - David Hill, Monterey Bay Whalewatch Center for Whale Research Learn more about killer whales in "L-Pod" |
The world's best air-breathing divers

One of the world's largest living mammals, the sperm whale is the deep-diving champion. The sperm whale dives down to nearly 2 km (one and a quarter miles) and stays under for over an hour on a regular basis. That puts it near food that includes colossal squid and the Patagonian tooth fish (Dissostichus eleginoides, also known as Chilean sea bass). At that dive range, sperm whales can access about a third of the world's ocean volume.
The much smaller beaked whale is a close second. Not much is known about these animals because they are rarely seen except when they've beached themselves.
Of the "smaller" mammals, elephant seals are the deepest divers.
Of the animals that are not mammals, a bird - the penguin, and a reptile - the sea turtle are also expert divers.
Comparison of dive statistics for air-breathing animals.
| Species | Maximum depth (meters) |
Maximum duration (minutes) |
Total oxygen stores (ml/kg> |
Hemoglobin (g/dl) |
Myoglobin (g/100g) |
Blood volume (ml/kg) |
| Humans Homo sapiens |
214 | 91 | 28 (20 | 14.5 | 0.3-0.4 | 70 |
| Leatherback turtle Dermochelys coriacea |
1230 | 67.3 | 27 | 15.6 | 4.9 | 77 |
| Bottlenose dolphin Tursiops truncatus |
390 | 8 | 36 | 14 | 3.3 | 71 |
| Emperor penguin Aptenodytes forsteri |
564 | 23.1 | 53 | 18 | 6.4 | 100 |
| Northern elephant seal Mirounga angustirostris |
1,581 | 119 | 97 | 25 | 6.5 | 250 |
| Beaked whale Ziphius cavirostris |
1,888 | 85 | unknown | unknown | 4.3 | unknown |
| Sperm Whale Physeter macrocephalus |
2,250 | 138 | 77 | 22 | 5.0 | 200 |
 Click to see reference list.
Updated: September 23, 2009.
Click to see reference list.
Updated: September 23, 2009.
Problems faced by human divers

Jessica diving in much shallower water than an elephant seal. Photographer - Kozue Shiomi.
Problems fall into three main categories -
- running out of oxygen - limits to the amount of oxygen our bodies carry
- the minimum oxygen levels at which our bodies remain functional
- negative behavior of gases

Animals like the elephant seal, whales, turtles, penguins, and dolphins breathe air into gas-filled lungs, just like us. They can only dive as far as a breath of air allows. So why can some animals dive much further before suffering ill consequences? Physiologists want to know how they do it and ecologists want to know why they do it.
Limited oxygen stores

Poor hypoxemic tolerance

Humans lose consciousness well before all the oxygen in their body is used up. Loss of consciousness while underwater leads to drowning and death. Elephant seals can function until their oxygen stores are nearly empty, allowing them to stay down longer on the same amount of oxygen.
Current research results

Jessica at her doctoral thesis defense.
 Free diver with monofin.
Photographer - Fathom.
Free diver with monofin.
Photographer - Fathom.When you try to hold your breath, that irresistible urge to take a breath is triggered by too high levels of carbon dioxide (CO2) gas rather than too low levels of oxygen (O2) gas. Sensitivity to elevated levels of CO2 before running out of O2 provides a safety cushion. It tells us that taking a breath is our highest priority.
CO2 is produced as a byproduct of respiration as your body metabolizes O2. During a breath hold dive, CO2 accumulates in your system unless you exhale.
Sometimes expert divers want to delay the signal from CO2 so they can stay under longer. By hyperventilating before diving, they lower their pre-dive CO2 load relative to O2. But sometimes they run out of oxygen before the CO2 trigger occurs and experience shallow water blackout that leads to drowning and death.
Decompression sickness

We tend to think of crushing pressures in the deep sea. We know that the downward pressure exerted by even one meter thickness of overlying water is more than that exerted by the entire columnn of air above sea level. Yet we also see delicate animals living at all depths in the sea without being crushed. Since water hardly compresses at all compared tp highly compressible gases such as air, animals that are filled with water do not experience the same problems faced by animals that are partly filled with air.
At high pressure, air compresses and changes from a gas into a liquid and it tends to dissolve into liquids such as water or blood. The reverse happens when pressure is reduced, liquefied gas turns back into a gas and expands significantly. Gases that were dissolved in water or blood come back out, sometimes in inopportune places.
What's a caisson?
A caisson† is a watertight structure that allows construction, mining and other work to go on inside. Caissons are used to build footings for bridges at the bottom of a river. Air that is pumped into a caisson structure at higher than normal pressure maintains the structure and keeps water out. Workers and the designer who built the foundations of the Brooklyn Bridge suffered many ailments when they exited the workplace. Modern health and safety regulations based on new knowledge protect people who work in caissons today.
• The Brooklyn Bridge, Harpers 1883
• Caisson disease during the construction of the Eads and Brooklyn Bridges: A review
† The term caisson has other meanings as well.
Avoiding decompression sickness in space
• NASA: Going out for a walk• NASA: Pass the S'mores Please! Station Crew 'Camps Out'
When someone dives and the surrounding pressure increases, the lungs compress as air compresses. Gases also dissolve into the body. When the pressure is reduced, the process reverses, gases in the lung expand and dissolved gas comes out of the body. When humans try to surface too fast, gas bubbles can form in blood vessels, joints and other unfortunate places rather than in the lungs. Gas bubbles in those places create immediate and delayed problems that can cause permanent damage as well as death. Thus divers learn the importance of a slow accent and that if needed to take a break just below the surface.
Not just diving, but any activity that involves a rapid drop in pressure can cause similar problems. Flying in an unpressurized aircraft and stepping into outer space are two such situations.
Elephant seals use several strategies to avoid decompression sickness. They don't carry compressible gas to the deep in the first place, so it's not there to reappear as they surface. They lack some of sinuses that house air in a human head. They expel air before diving by collapsing lungs encased in flexible rather than rigid ribs.
Beaked whales may suffer decompression sickness

Beaked whales comprise 21 species of rarely seen and little understood marine mammals that look somewhat like overgrown dolphins. They've been in the news because of mass strandings.
 Cuvier's beaked whale - Ziphius cavirostris.
Image from the Smithsonian Institution.
Cuvier's beaked whale - Ziphius cavirostris.
Image from the Smithsonian Institution.
Learn more about beaked whales • Diving with Beaked Whales - video
• Smithsonian Institution
• NOAA
• BBC News
Fifteen of these animals beached themselves and died in the Canary Islands in 2002 after a military sonar test. Because some strandings are coincident with sonar tests, their effect on whales is a concern. But the cause-and-effect relationships are still unclear. Whales of other types have been known to beach themselves in the absence of sonar too.
The beaked whale strandings in the Canary Islands raised a unique concern, as necropsies revealed injured tissues in blood vessels and vital organs similar to that which could be caused by decompression sickness. Traditionally, diving animals have not been thought to suffer from decompression sickness like humans. The question remains whether or not beaked whales could be susceptible to this problem, and many researchers are currently investigating the science behind this issue. [33]
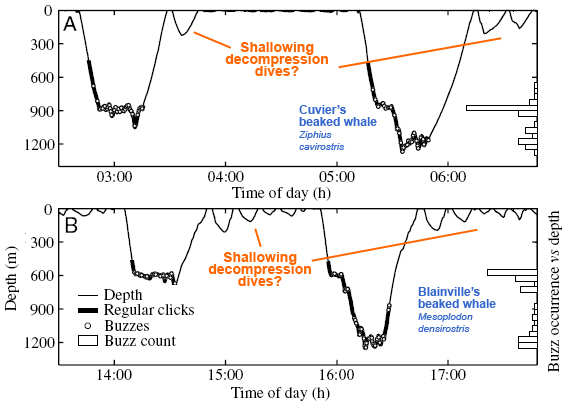 Diving pattern of the beaked whale
Diving pattern of the beaked whale
The dive pattern of the beaked whale differs from the elephant seal pattern. Both animals descend rapidly, but the beaked whale comes up to the surface and then takes several shallow dives before going down again [34]. This process may help them avoid decompression sickness in the same way that a pattern of decompression stops help human divers ascend safely. Noises that keep the animal from making necessary decompression dives may cause it to become sick and die.
Some scientists speculate that sonar might trigger the formation of bubbles in the bodies of these animals directly. The more we know about individual species of animals, the more we can fine-tune our activities. Knowing more might prevent us from doing harm in one place, while letting us operate freely in others.
Nitrogen narcosis

When the concentration of nitrogen becomes too rich, at high pressure at depth, humans experience nitrogen narcosis or rapture of the deep. Effects resemble alcohol intoxication and include disorientation, very poor judgment, and euphoria. Different mixes of dive gases offer advantages, but also carry different risks. Any effect that impairs judgment is extremely hazardous while diving.
All of these problems are complex and not entirely understood. Diving provides a rare opportunity to experience an alien world and lifestyle, but it can be unforgiving when you're a human rather than an elephant seal.